連続について
生命は生と死の二分法によって規定され、生命の設計図はDNAの塩基配列であり、どちらも離散的な1で構成されていると見なし得る。さらに神経細胞の活動電位も全か無の法則に従う。こうして考えると、生命はデジタル信号を好むように思われる。特にウイルスなどは、細胞が無くDNAまたはRNAと生死だけであるように見え、コンピューターウイルスと同型に思われる。そうであるのに脳は空間と時間を連続的なものとして認識し、物体も連続的なものとして認識する。そうすると連続とは何なのか、この問題に関して西洋で主流となったのは、アリストテレスの考えである(1)。アリストテレスは、連続の最も大きな特徴はいくらでも分割できることであると述べて、さらに連続的なものは、分割不可能な最小の要素から出来ているのではないと述べている。アリストテレスによると時間は瞬間から出来ているのではなく、線は点から出来ているのではないという事になる。むしろ点と線と面の関係で述べたように、点は線の切断によって生じるのであるから、瞬間は時間の切断と考えるべきだろう。こうして考えると生命及び神経にとっての認識の基本要素である「1」は、連続的なものの基本要素ではない事になる。このように通常の神経系と異なる認識方法は、何のために存在するのか、果たして連続という認識方法は利点があるのか。この問題を中心に考察する。
連続というのは時間と空間に共通する性質であるが、時間と空間に同時に関係する現象は運動である。動物にとって運動は非常に重要であり、運動のために神経系が発生したとも考えられる。人間は運動を連続的なものとして認識する。ところが1983年に、両則性の側頭葉と後頭葉の中間部分の脳梗塞で、運動視が特異的に障害された症例が報告された(2)。患者は立体視は問題なく、色も識別でき、視力も良く、簡単に物体を見分けられた。ところが患者は三次元的な動きが全く感知できないと訴えていた。例えば紅茶やコーヒーをカップに注ごうとしても、液体が氷河のように固まって見えるため上手くいかない。その他にも連続的な動きは全く認識でいなかった。この症例での病巣は、サルにおいては運動視の中枢と考えられているMT野に相当する。MT野のニューロンはV1野、V2野、V3野からの入力を受けている(3)が、全て離散的な活動電位である。MT野のニューロンは離散的な活動電位の入力から、連続的な動きを創り出すものと考えられる。網膜で光を受容して、神経節細胞の活動電位が発生してからMT野に至るまでの間、どこにも連続的な要素はない。活動電位が発生する以前の段階についても、良く研究されている桿体について考察する。最も鋭敏な反応では1個の光子を検出し、1個のロドプシンが構造変化する。この反応では、原理的には活性型ロドプシンの個数は自然数に換算できる。これは特定の視細胞で起こるので、網膜上の位置もデジタル的に決定される。また、この反応は非常に短時間で起こるので時刻も特定される。すなわち光の受容の第一段階ではは、強度、位置、時刻の全てがデジタル的に決定されている。次に視野内における小さな物体の移動を考えてみる。神経系は第一段階では、特定の時刻における物体の視野内の位置を、とびとびの離散的なデータとして認識するだけである。人間の神経系はこのデータを基に計算によって連続的な運動を創り出すのである。これは複雑な過程であるので、自然淘汰における利点があるはずである。次にその点を考察する。
私は高等脊椎動物が生存競争に打ち勝つには、運動を連続的なものとして認識する必要があったのではないかと考えている。それは大型哺乳類や鷲や鷹のように体の大きい動物は、網膜の視細胞で光を認識してから運動に至るまで、多くのシナプスと長い軸索を経なくてはいけないので、どうしても時間がかかるという不利がある。それに対して昆虫は体の大きさから考えて、視覚認知から反応までの時間が短くて済むと考えられる。さらに反応速度という点だけを考えれば、単細胞のユーグレナのように細胞内に光受容体があれば、光を受けてすぐに反応できる。反応時間の差を補うための一つの解答として、私は高等脊椎動物が物体または他の動物の運動を予測するのに、ニュートン力学を利用すると考えている。光受容体が光を受けてから体が動くまでの反応速度だけでは、信号を伝達する距離の短い昆虫が有利である。そのため高等脊椎動物が昆虫を捕らえようとする場合、一匹の昆虫の飛ぶ軌道をニュートン力学を用いて予測し、先回りしていると思われる。このような計算の具体例として、現代の文明社会における人間であれば、野球について考えるとわかりやすい。ピッチャーがボールを投げてすぐにバッターのスイングは始まる。つまりピッチャーの投げたボールの軌道を予測することによって、バッターは打つことが出来る。また外野手はバッターが打ってから、かなり早い時期に走り始める。つまり外野手は打球の落下点を予測してキャッチする。このように人間の神経系は、打球の放物線の軌道に、風の影響なども加味して計算する能力を持っている。これらの計算にはニュートン力学が必要であり、ニュートン力学の前提条件は時間と空間および運動の連続性である。つまり人間のような高等脊椎動物は、時間と空間を連続的と認知して、ニュートン力学を用いた計算を行い、生存競争に打ち勝ってきたと考えられる。この方法は、反応速度を超越した利点があると考えられる。このように考えると、運動の連続性こそが本質的であり、そのために時間と空間の連続性が生じたと考えられる。

図1にボールが壁の間を往復するアニメーションを示した。これは見た目にはボールが連続的に移動しているように見える。しかし実際には1/10秒間隔で静止画を入れ替えているだけである。つまり連続的な運動は不連続な動きから脳内で創作されたものである。
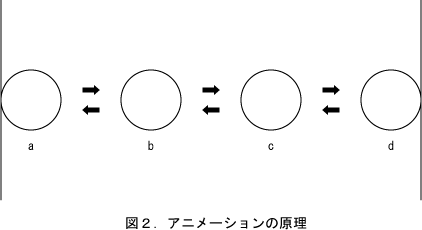
図2においてアニメーションの原理を示した。1個のボールがa地点からd地点までの4カ所にある4枚の絵を用意し、一定の時間間隔で順番に入れ替えていくと、ボールが連続的に移動しているかに見える。最初はそのようにして図1を作ったのであるが、ややぎこちなく見えるので、片道7枚に増やした上で往復運動にして図1とし、結果的には往復12枚のアニメーションとなった。このようにアニメーションは静止画を入れ替えているだけであり、映画でも1秒間に24枚の写真を入れ替えているだけである。つまり人間は脳内において、離散的な少しの情報から連続的な動きを作り出している。この事実については、D.D.Hoffmanの「視覚の文法」(4)に多くの例が説明されている。
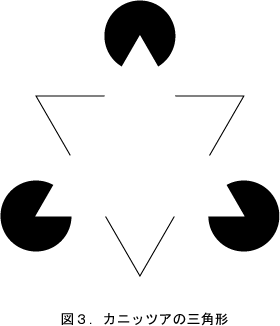
次に「視覚の文法」では運動以外にも、脳内で連続を創り出す例として、主観的輪郭も多く取り上げられている。最もよく知られているのが、図3に示したカニッツアの三角形である。ここで中央に明るい白い三角形が浮かび上がって見える。その輪郭を主観的輪郭というが、実際には線は存在せず、これこそが全く脳で作られたエッジである。そこから得られる結論は、連続は脳で作り出されたということになる。人間には物体のエッジを連続的なものとして認識しようとする傾向があり、見える部分だけでなく、見えない部分まで物体のエッジをつなごうとする。また三角形や円を好む傾向もある。そうすると、この図形は前面に明るい三角形があり、後方に三角形と三つの円があるとすると、一番安定した解釈となる。
こうして連続性は脳内で形成される。それに対して脳に入る信号も、脳から出る信号も離散的である。これを前提として連続の役割について考えてみる。人間の神経系は複雑であり体も大きいので、神経の軸索も長くなりシナプスの数も多くなる。このため神経系への入力に反応するだけでは、速度的に不利な場合が生じる。特に動く物体や動物を相手にした場合は、反応が間に合わない。そこで予測を用いる。これを原始人が狩りをする場合を想定して考えてみる。最初に視覚的に獲物の位置を認識するのであるが、視神経から脳へ入る信号は全て活動電位であるので、デジタル的に認識するしかない。それを連続的なデータに変換することによって、ニュートン力学を成立させる。ニュートン力学の前提は時間と空間の連続性であるので、連続的な世界を脳内に創作するのである。その上でニュートン力学を用いて獲物の運動を計算し、未来の獲物の位置を予測する。そして予測に基づいて槍を投げて獲物を倒すのである。最後の槍を投げる段階では、連続的なデータを再びデジタル信号に直して筋肉に活動電位を送る必要がある。このように脳内では連続的な認識は有効であるが、現実に行動するときはデジタル化しなくてはならない。ただしこれは世界がデジタル的であるという意味ではない。ただ人間の神経系が外界をデジタル化して認識し、外界への出力もデジタル信号であるというだけである。デジタル化は人間の感覚器官が行っているのであって、世界そのものがデジタルであることを意味しない。むしろ世界はアナログ的であり、時間と空間は連続であると考える方が一般的である。しかし、人間の感覚器官が感覚をデジタル化するため、世界が連続的であるとしても確認しようがない。結局のところ、世界そのものはデジタルであるかもしれないし、アナログであるかもしれず真の解答は不明である。もしかすると、この二つの認識方法だけでは、真実の世界を認識するには不十分なのかもしれない。
参考文献
(1) Aristotle:Physics,Oxford University Press(1996)
(2) J.Zihl, D.von Cramon, N.Mai: Brain, 106, 313(1983)
(3) 福田淳・佐藤宏道:脳と視覚,共立出版(2002)
(4) D.D.ホフマン:視覚の文法,原淳子・望月弘子訳,紀伊國屋書店(2003)
前のページ | 目次 | 次のページ